Intermittent Fasting Ameliorated High-Fat Diet-Induced Memory Impairment in Rats via Reducing Oxidative Stress and Glial Fibrillary Acidic Protein Expression in Brain

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Experimental Design
2.3. Sampling
2.4. Assessment of Behavioral Responses
2.5. Assessment of Motor Function
Open Field Test
2.6. Assessment of Short-Term Spatial Memory
Y-Maze
2.7. Biochemical Investigations
2.8. Measurement of Brain Tissues Malondialdehyde and Reduced Glutathione Concentrations
2.9. Histopathological Examination
2.10. Immunohistochemical Investigations
2.11. Statistical Analysis
3. Results
3.1. Intermittent Fasting Ameliorated HFD Increased Rats’ Body Weight
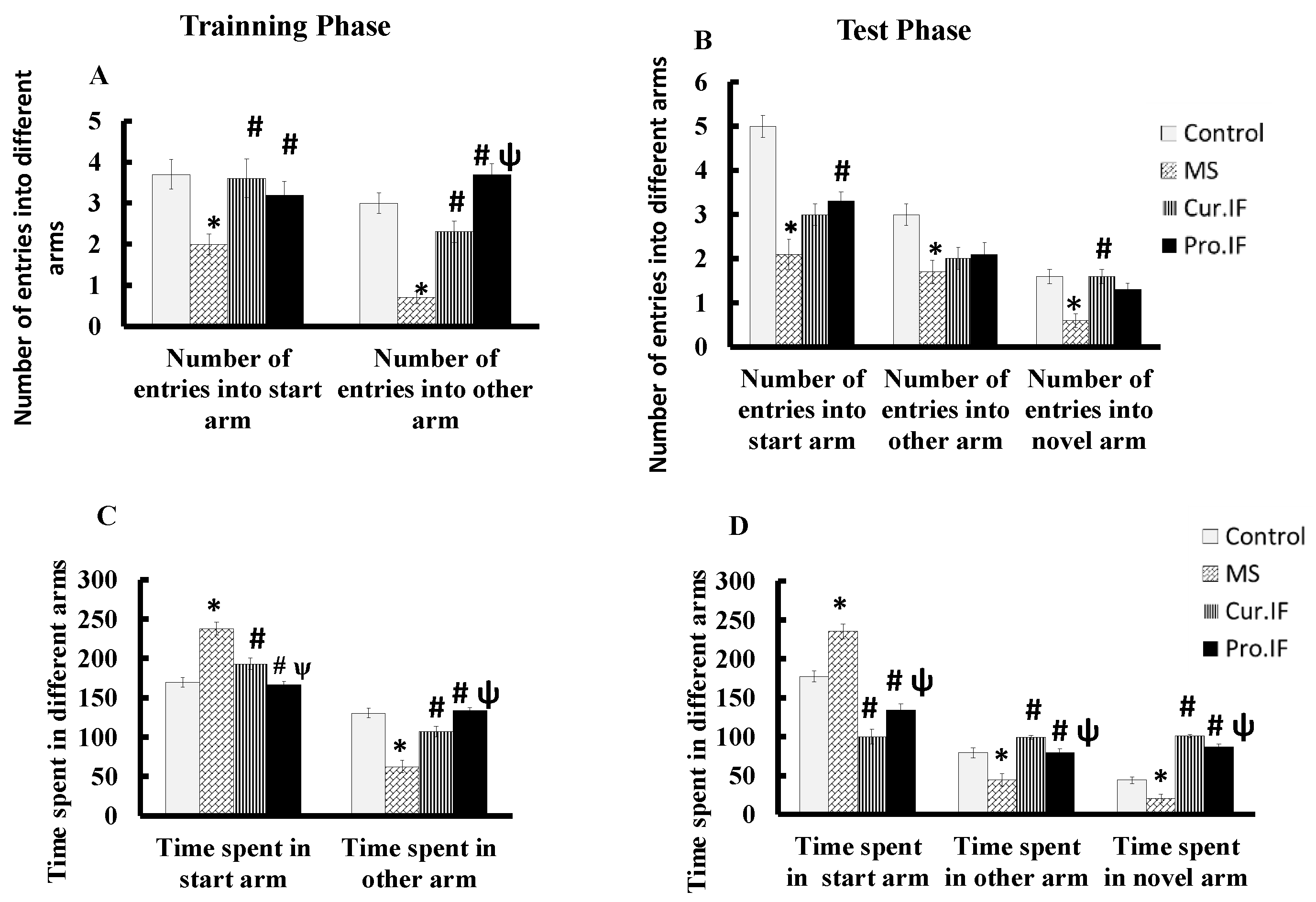
3.2. Intermittent Fasting Improved the Behaviors of HFD Fed Rats
3.2.1. Open Field
3.2.2. Y-Maze
3.3. Intermittent Fasting Ameliorated HFD Altered Serum Lipid Profile in Rats
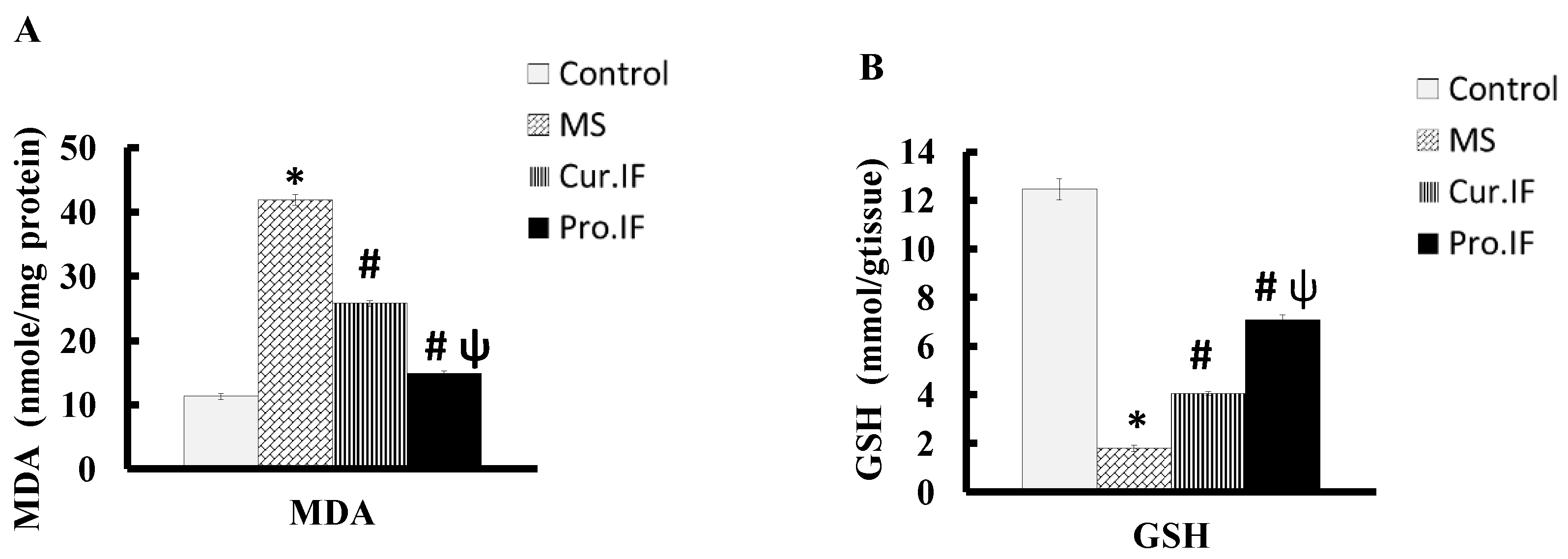
3.4. Intermittent Fasting-Modulated HFD Altered Malondialdehyde and Reduced Glutathione Concentrations in the Brain Tissues
3.5. Intermittent Fasting-Ameliorated HFD Induced Histopathological Changes in Brain Tissues of Rats
3.6. Intermittent Fasting Reduced HFD Increased GFAP Protein Expression in Brain Tissues of Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lasker, S.; Rahman, M.M.; Parvez, F.; Zamila, M.; Miah, P.; Nahar, K.; Kabir, F.; Sharmin, S.B.; Subhan, N.; Ahsan, G.U.; et al. High-fat diet-induced metabolic syndrome and oxidative stress in obese rats are ameliorated by yogurt supplementation. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.R.; Haley-Zitlin, V.; Rosenberger, D.S.; Granholm, A.-C. Damaging effects of a high-fat diet to the brain and cognition: A review of proposed mechanisms. Nutritional 2014, 17, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Spencer, S.J.; D’Angelo, H.; Soch, A.; Watkins, L.R.; Maier, S.F.; Barrientos, R.M. High-fat diet and aging interact to produce neuroinflammation and impair hippocampal-and amygdalar-dependent memory. Neurobiol. Aging 2017, 58, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Forti, P.; Pisacane, N.; Rietti, E.; Lucicesare, A.; Olivelli, V.; Mariani, E.; Mecocci, P.; Ravaglia, G. Metabolic syndrome and risk of dementia in older adults. J. Am. Geriatr. Soc. 2010, 58, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Balasubramanian, P.; Weerasekara, Y.K. Nutrition modulation of human aging: The calorie restriction paradigm. Mol. Cell. Endocrinol. 2017, 455, 148–157. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, M.; Chen, Y.; Yang, Y.; Zhang, J.-J. Postoperative intermittent fasting prevents hippocampal oxidative stress and memory deficits in a rat model of chronic cerebral hypoperfusion. Eur. J. Nutr. 2019, 58, 423–432. [Google Scholar] [CrossRef]
- Qin, W.; Chachich, M.; Lane, M.; Roth, G.; Bryant, M.; de Cabo, R.; Ottinger, M.A.; Mattison, J.; Ingram, D.; Gandy, S.J. Calorie restriction attenuates Alzheimer’s disease type brain amyloidosis in Squirrel monkeys (Saimiri sciureus). J. Alzheimer’s Dis. 2006, 10, 417–422. [Google Scholar] [CrossRef]
- Singh, R.; Lakhanpal, D.; Kumar, S.; Sharma, S.; Kataria, H.; Kaur, M.; Kaur, G. Late-onset intermittent fasting dietary restriction as a potential intervention to retard age-associated brain function impairments in male rats. Age 2012, 34, 917–933. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Manchanda, S.; Kaur, T.; Kumar, S.; Lakhanpal, D.; Lakhman, S.S.; Kaur, G. Middle age onset short-term intermittent fasting dietary restriction prevents brain function impairments in male Wistar rats. Biogerontology 2015, 16, 775–788. [Google Scholar] [CrossRef]
- Morgan, T.; Wong, A.; Finch, C. Anti-inflammatory mechanisms of dietary restriction in slowing aging processes. In Mechanisms of Dietary Restriction in Aging and Disease; Karger Publishers: Los Angeles, CA, USA, 2007; Volume 35, pp. 83–97. [Google Scholar]
- Wan, R.; Ahmet, I.; Brown, M.; Cheng, A.; Kamimura, N.; Talan, M.; Mattson, M.P. Cardioprotective effect of intermittent fasting is associated with an elevation of adiponectin levels in rats. J. Nutr. Biochem. 2010, 21, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Patterson, R.E.; Sears, D.D. Metabolic effects of intermittent fasting. Annu. Rev. Nutr. 2017, 37, 371–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Page, A.J.; Hatzinikolas, G.; Chen, M.; Wittert, G.A.; Heilbronn, L.K. Intermittent fasting improves glucose tolerance and promotes adipose tissue remodeling in male mice fed a high-fat diet. Endocrinology 2019, 160, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Wang, Z.; Zuo, Z.J. Chronic intermittent fasting improves cognitive functions and brain structures in mice. PLoS ONE 2013, 8, e66069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albus, U. Guide for the Care and Use of Laboratory Animals, 8th ed.; SAGE Publications Sage: London, UK, 2012. [Google Scholar]
- Ferragud, A.; Velázquez-Sánchez, C.; Abdullatif, A.A.; Sabino, V.; Cottone, P. Withdrawal from Extended, Intermittent Access to A Highly Palatable Diet Impairs Hippocampal Memory Function and Neurogenesis: Effects of Memantine. Nutrients 2020, 12, 1520. [Google Scholar] [CrossRef]
- He, D.; Mustafi, D.; Fan, X.; Fernandez, S.; Markiewicz, E.; Zamora, M.; Mueller, J.; Sachleben, J.R.; Brady, M.J.; Conzen, S.D.; et al. Magnetic resonance spectroscopy detects differential lipid composition in mammary glands on low fat, high animal fat versus high fructose diets. PLoS ONE 2018, 13, e0190929. [Google Scholar] [CrossRef] [Green Version]
- Matheus, F.C.; Rial, D.; Real, J.I.; Lemos, C.; Ben, J.; Guaita, G.O.; Pita, I.R.; Sequeira, A.C.; Pereira, F.C.; Walz, R.; et al. Decreased synaptic plasticity in the medial prefrontal cortex underlies short-term memory deficits in 6-OHDA-lesioned rats. Behav. Brain Res. 2016, 301, 43–54. [Google Scholar] [CrossRef]
- Soares, E.; Prediger, R.D.; Nunes, S.; Castro, A.A.; Viana, S.D.; Lemos, C.; De Souza, C.M.; Agostinho, P.; Cunha, R.A.; Carvalho, E.; et al. Spatial memory impairments in a prediabetic rat model. Neuroscience 2013, 250, 565–577. [Google Scholar] [CrossRef]
- DeLong, D.M.; DeLong, E.R.; Wood, P.D.; Lippel, K.; Rifkind, B.M. A comparison of methods for the estimation of plasma low-and very low-density lipoprotein cholesterol: The Lipid Research Clinics Prevalence Study. JAMA 1986, 256, 2372–2377. [Google Scholar] [CrossRef]
- Wilson, P.W.; Abbott, R.D.; Garrison, R.J.; Castelli, W.P. Estimation of very-low-density lipoprotein cholesterol from data on triglyceride concentration in plasma. Clin. Chem. 1981, 27, 2008–2010. [Google Scholar] [CrossRef]
- Draper, H.; Squires, E.; Mahmoodi, H.; Wu, J.; Agarwal, S.; Hadley, M. A comparative evaluation of thiobarbituric acid methods for the determination of malondialdehyde in biological materials. Free Radic. Biol. Med. 1993, 15, 353–363. [Google Scholar] [CrossRef]
- Beutler, E.; Dubon, O.; Kelly, B.M. Improved method for the determination of blood glutathiones. J. Lab. Clin. Med. 1963, 61, 882–890. [Google Scholar] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques, 6th ed.; Churchill Livingstone, Elsevier Health Sciences: Beijing, China, 2008. [Google Scholar]
- Kiernan, J.A. Histological and histochemical methods: Theory and practice. Schock 1999, 12, 479. [Google Scholar]
- Pirker, R.; Pereira, J.R.; Von Pawel, J.; Krzakowski, M.; Ramlau, R.; Park, K.; De Marinis, F.; Eberhardt, W.E.; Paz-Ares, L.; Störkel, S.; et al. EGFR expression as a predictor of survival for first-line chemotherapy plus cetuximab in patients with advanced non-small-cell lung cancer: Analysis of data from the phase 3 FLEX study. Lancet Oncol. 2012, 13, 33–42. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Abd Eldaim, M.A.; Ibrahim, F.M.; Orabi, S.H.; Hassan, A.; El Sabagh, H.S. l-Carnitine-induced amelioration of HFD-induced hepatic dysfunction is accompanied by a reduction in hepatic TNF-α and TGF-β1. Biochem. Cell Biol. 2018, 96, 713–725. [Google Scholar] [CrossRef]
- Orabi, S.H.; Al-Sabbagh, E.S.; Khalifa, H.K.; Mohamed, M.A.E.G.; Elhamouly, M.; Gad-Allah, S.M.; Abdel-Daim, M.M.; Eldaim, M.A.A. Commiphora myrrha Resin Alcoholic Extract Ameliorates High Fat Diet Induced Obesity via Regulation of UCP1 and Adiponectin Proteins Expression in Rats. Nutrients 2020, 12, 803. [Google Scholar] [CrossRef] [Green Version]
- Seibenhener, M.L.; Wooten, M.C. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.F.; Xie, Z.X.; Qiao, Y.; Li, L.R.; Cheng, X.R.; Tang, X.; Shi, Y.H.; Le, G.W. Differential effects of quercetin on hippocampus-dependent learning and memory in mice fed with different diets related with oxidative stress. Physiol. Behav. 2015, 138, 325–331. [Google Scholar] [CrossRef]
- Mielke, J.G.; Nicolitch, K.; Avellaneda, V.; Earlam, K.; Ahuja, T.; Mealing, G.; Messier, C. Longitudinal study of the effects of a high-fat diet on glucose regulation, hippocampal function, and cerebral insulin sensitivity in C57BL/6 mice. Behav. Brain Res. 2006, 175, 374–382. [Google Scholar] [CrossRef]
- Cordner, Z.A.; Tamashiro, K.L. Effects of high-fat diet exposure on learning & memory. Physiol. Behav. 2015, 152, 363–371. [Google Scholar]
- Kothari, V.; Luo, Y.; Tornabene, T.; O’Neill, A.M.; Greene, M.W.; Geetha, T.; Babu, J.R. High fat diet induces brain insulin resistance and cognitive impairment in mice. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2017, 1863, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Luchsinger, J.A.; Tang, M.-X.; Shea, S.; Mayeux, R.J. Caloric intake and the risk of Alzheimer disease. Arch. Neurol. 2002, 59, 1258–1263. [Google Scholar] [CrossRef] [PubMed]
- Valladolid-Acebes, I.; Stucchi, P.; Cano, V.; Fernández-Alfonso, M.S.; Merino, B.; Gil-Ortega, M.; Fole, A.; Morales, L.; Ruiz-Gayo, M.; Del Olmo, N. High-fat diets impair spatial learning in the radial-arm maze in mice. Neurobiol. Learn. 2011, 95, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Beilharz, J.E.; Maniam, J.; Morris, M.J. Short exposure to a diet rich in both fat and sugar or sugar alone impairs place, but not object recognition memory in rats. Brain Behav. Immun. 2014, 37, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Tran, D.M.; Westbrook, R.F. A high-fat high-sugar diet-induced impairment in place-recognition memory is reversible and training-dependent. Appetite 2017, 110, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Bondan, E.F.; Cardoso, C.V.; Martins, M.D.F.M.; Otton, R. Prejuízos de memória e expressão aumentada de GFAP em astrócitos hipocampais após dieta hipercalórica em ratos. Arquivos de Neuro-Psiquiatria 2019, 77, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Rebrin, I.; Forster, M.J.; Sohal, R.S. Effects of age and caloric intake on glutathione redox state in different brain regions of C57BL/6 and DBA/2 mice. Brain Res. 2007, 1127, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-H.; Ouyang, C.; Ding, Q.; Song, J.; Cao, W.; Mao, L. A moderate low-carbohydrate low-calorie diet improves lipid profile, insulin sensitivity and adiponectin expression in rats. Nutrients 2015, 7, 4724–4738. [Google Scholar] [CrossRef] [Green Version]
- Das, B.; Mishra, T. Role of HDL-C in health and disease. J. Indian Acad. Clin. Med. 2012, 13, 218–222. [Google Scholar]
- Marbut, M.M.; Al-Najjar, H.A.D.S.; Mustafa, A. Effect of Ramadan fasting on some physiological parameters. Med. J. Tikrit Univ. 2005, 2, 6–8. [Google Scholar]
- Nurmasitoh, T.; Utami, S.Y.; Kusumawardani, E.; Najmuddin, A.A.; Fidianingsih, I. Intermittent fasting decreases oxidative stress parameters in Wistar rats (Rattus norvegicus). Univ. Med. 2018, 37, 31–38. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Constituents | Control Group |
|---|---|
| Fat | 7–10% |
| Carbohydrates | 68–70% |
| Protein | 18–20% |
| Vitamins and minerals | 1–2% |
| Kcal/100 g/day | 341 |
| Diet Constituents | HFD |
|---|---|
| Fat | 30% |
| Carbohydrates | 50–52% |
| Protein | 18–20% |
| Vitamins and minerals | 1–2% |
| Kcal/100 g/day | 530 |
| Metabolite | Control | HFD | HFD & Cur IF | HFD & Pro IF |
|---|---|---|---|---|
| Cholesterol (mg/dL) | 119.60 ± 1.88 | 277.10 ± 1.84 * | 174.50 ± 2.37 # | 129 ± 1.24 #ψ |
| Triglycerides (mg/dL) | 122 ± 3.54 | 203.00 ± 7.20 * | 174.70 ± 2.07 # | 134.70 ± 2.54 #ψ |
| LDL (mg/dL) | 43.60 ± 3.77 | 209.78 ± 2.22 * | 109.06 ± 2.18 # | 70.16 ± 1.94 #ψ |
| HDL(mg/dL) | 51.60 ± 2.52 | 26.70 ± 0.97 * | 31.50 ± 1.82 | 39.00 ± 1.52 #ψ |
| VLDL (mg/dL) | 24.40 ± 0.70 | 40.62 ± 0.63 * | 34.94 ± 0.41 # | 26.94 ± 0.50 #ψ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hazzaa, S.M.; Eldaim, M.A.A.; Fouda, A.A.; Mohamed, A.S.E.D.; Soliman, M.M.; Elgizawy, E.I. Intermittent Fasting Ameliorated High-Fat Diet-Induced Memory Impairment in Rats via Reducing Oxidative Stress and Glial Fibrillary Acidic Protein Expression in Brain. Nutrients 2021, 13, 10. https://doi.org/10.3390/nu13010010
Hazzaa SM, Eldaim MAA, Fouda AA, Mohamed ASED, Soliman MM, Elgizawy EI. Intermittent Fasting Ameliorated High-Fat Diet-Induced Memory Impairment in Rats via Reducing Oxidative Stress and Glial Fibrillary Acidic Protein Expression in Brain. Nutrients. 2021; 13(1):10. https://doi.org/10.3390/nu13010010
Chicago/Turabian StyleHazzaa, Suzan M., Mabrouk A. Abd Eldaim, Amira A. Fouda, Asmaa Shams El Dein Mohamed, Mohamed Mohamed Soliman, and Eman I. Elgizawy. 2021. "Intermittent Fasting Ameliorated High-Fat Diet-Induced Memory Impairment in Rats via Reducing Oxidative Stress and Glial Fibrillary Acidic Protein Expression in Brain" Nutrients 13, no. 1: 10. https://doi.org/10.3390/nu13010010